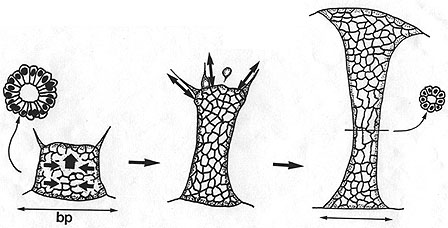
A schematic diagram depicting the forcers acting during archenteron elongation in the sea urchin embryo. Early during secondary invagination, active rearrangement of cells in the wall of the archenteron occurs, followed by secondary mesenchyme-dependent elongation, mediated by the traction produced by SMCs. Image by Jeff Hardin, Univ. of Wisconsin.
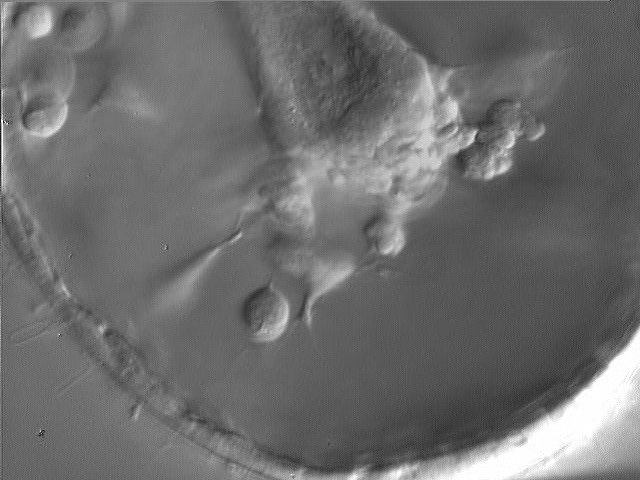
Secondary mesenchyme cells (SMCs) constantly extend and retract filopodia as the archenteron elongates (click on the movie thumbnail on the left at the bottom of this page for an example). Filopodia clearly exert tension where they attach, pulling out what Tryggve Gustafson called "cones of attachment". When a filopodium detaches, such cones rapidly snap back, indicating that the filopodia were exerting tension where they were attached.
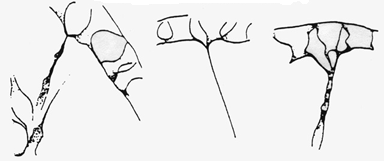
Top: Tracings of individual filopodia, pulling out "cones of attachment". Original drawing by Tryggve Gustafson; reproduced from Hardin (1996), with permission. Bottom: frame from a movie showing a cone of attachment (arrow) produced by a SMC in L. pictus. Image by Jeff Hardin, Univ. of Wisconsin.
To see movies of SMCs and cones of attachment, click on the thumbnails.
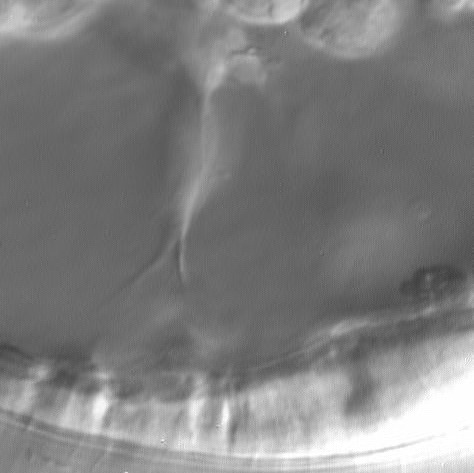 |
 |
|
Cones of attachment
(1.2 Mb)
|
Late archenteron
tip (1.3 Mb)
|
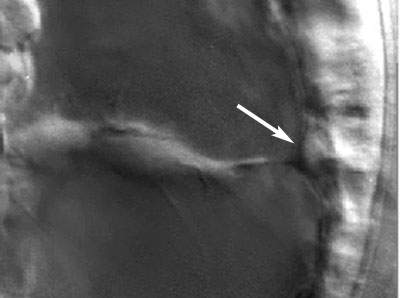
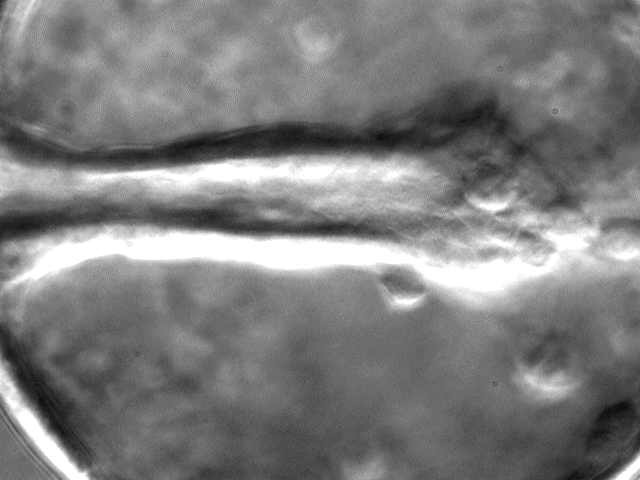
If SMCs are important near the end of gastrulation, then we would predict that pulling by SMCs would be more substantial late in gastrulation. In fact, the archenteron displays "necking" late in gastrulation, a phenomenon well known to engineers as a characteristic of plastic deformation (sort of like pulling salt water taffy).

Frame from a movie showing "necking" of the archenteron, likely produced by SMCs, in L. pictus. Image by Jeff Hardin, Univ. of Wisconsin.
Click on the thumbnail to see a time-lapse movie that demonstrates this necking behavior.
 |
|
Completion of
gastrulation in L. pictus (1.0 Mb)
|