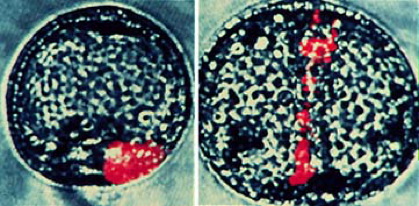
L. pictus gastrulae containing clones of cells that have incorporated into the vegetal plate. At the onset of gatrulation, such clones are a contiguous block of tissue (left). Image by Jeff Hardin, Univ. of Wisconsin.
Although we now know that convergent extension occurs, it has been difficult to study it in vivo. I used the species Eucidaris tribuloides (a pencil urchin from the Caribbean) to study this process. The embryos of this species are incredibly transparent, so convergent extension can be visualized directly.
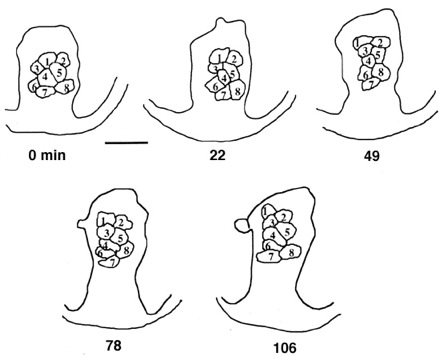
Tracings from a time-lapse movie of a Eucidaris tribuloides gastrula. the cells numbered 1-8 visibly change position during the period of the movie (time shown is in minutes). Image by Jeff Hardin, Univ. of Wisconsin.
Since the secondary mesenchyme cells (SMCs) in Eucidaris gastrulae never attach to the animal pole, their SMCs can't be pulling upward to any appreciable extent, providing a "natural experiment" for studying active convergent extension.
More direct evidence that the cells in the archenteron can rearrange without the pulling forces of SMCs came from other experiments.
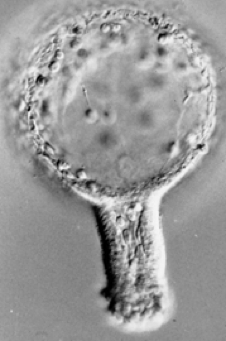
To test whether secondary mesenchyme cells are actually needed for archenteron elongation, I performed several experiments as a graduate student (!). First, it is possible to produce "exogastrulae", in which the archenteron evaginates, rather than invaginating (my wife, Susie, affectionately calls such embryos "outies"). In this case, the archenteron does elongate, although it is roughly 2/3 of its normal final length. In fact, the same rearrangement processes that occur in normal archenteron seem to occur in such embryos.
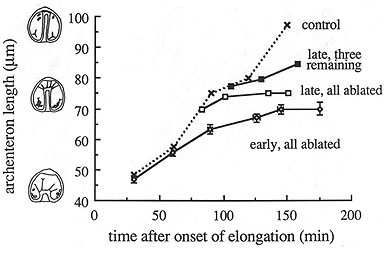
A direct test to determine whether secondary mesenchyme cells are actually needed for archenteron elongation involves killing them using a laser microbeam. I performed this experiment (in 1988!), and found that the archenteron can elongate to 2/3 of its normal, final length in the absence of functional SMC filopodia. This shows that the archenteron can elongate on its own, but that it requires intact SMCs for complete elongation.
This result can be quantified, by correlating the rate of migration with the number of intact SMCs, as shown in the graph below. This graph shows that there is a quantitative relationship between rate of extension in late gastrulae and the number of intact SMCs. Remarkably, as few as three intact SMCs are sufficient for continued elongation, although elongation occurs more slowly.